From: Treseder KK, Lennon JT. 2015. Fungal traits that drive ecosystem dynamics on land. Microbiology and Molecular Biology Reviews 79:243-262.

Fungal cell walls provide protection from desiccation, freeze-thaw damage, and other environmental stresses (Bowman and Free 2006, Latgé 2007). Most fungal taxa construct cell walls with chitin (James et al. 2013); some can incorporate β1,3-glucan as well (Latge and Calderone 2005, Xie and Lipke 2010). β1,3-glucan is a carbohydrate that forms cross-linkages with chitin and other components (Cabib 2009), improving the strength and integrity of the cell wall (Kollar et al. 1997). In fact, mutants of Saccharomyces cerevisiae that lack the ability to synthesize β1,3-glucan are about 5-fold more sensitive to drought stress than wild type (Shima et al. 2008). β1,3-glucan can constitute as much as 55% of the dry weight of the fungal cell wall (Klis 1994). Moreover, it is highly polymerized, hydrophobic, and acid- and alkali-insoluble when cross-linked with chitin (Klis 1994), which could make it relatively resistant to decomposition. Although few studies to date have assessed turnover rates or standing stocks of β1,3-glucan in soils, it is worth investigating as a potentially significant component of microbial residues within ecosystems (Kogel-Knabner 2002). If so, β1,3-glucan could be a mechanism facilitating soil C storage in response to drought or other environmental stressors.
Fungal genes for β1,3-glucan synthesis
I have a question for readers: What other compounds produced by fungi might form recalcitrant C in the soil? I discuss melanin in another post. There must be others, though. If you have a suggestion, please leave a comment. Thanks!
http://mmbr.asm.org/content/79/2/243/F7.expansion.html
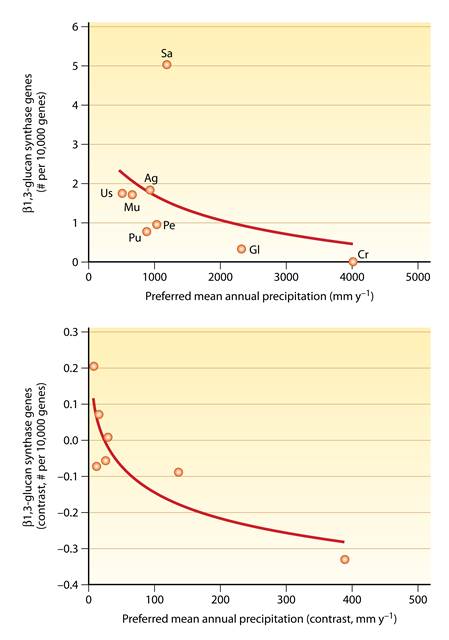
Since fungal phyla and subphyla varied in their genetic capacity for stress tolerance, we might expect their environmental distributions to covary accordingly, with stress-tolerators occupying harsher climates. In a recent large-scale study, Treseder et al. (2014) reported that ancient fungal phyla were relatively constrained to regions with higher precipitation levels, whereas younger phyla occurred in dry as well as wet ecosystems. The underlying physiological or morphological trait driving these differences in environmental preferences remained unknown. However, we found that the capacity to produce β1,3-glucan was linked to the preferred precipitation levels of fungi. For example, Cryptomycota, the oldest phylum, did not possess any known β1,3-glucan synthase genes. Correspondingly, they preferred wetter habitats, with average precipitation rates of 4,000 mm y-1. In contrast, the younger phyla/subphyla preferred drier sites, with the exception of Glomeromycota, which contained the lowest frequency of β1,3-glucan synthase genes among this group. It is possible that the capacity to produce β1,3-glucan may be an important trait allowing fungi to tolerate drought stresses typical of ecosystems with low rainfall.
β1,3-glucan synthase varies most at the subphylum level:

http://mmbr.asm.org/content/79/2/243/F3.expansion.html
Phylogenetic distribution of β1,3-glucan synthase genes:

http://mmbr.asm.org/content/79/2/243/F4.expansion.html
β1,3-glucan synthase genes are most abundant in yeasts.

http://mmbr.asm.org/content/79/2/243/F5.expansion.html
Bowman, S. M. and S. J. Free. 2006. The structure and synthesis of the fungal cell wall. Bioessays 28:799-808.
Cabib, E. 2009. Two novel techniques for determination of polysaccharide cross-links show that Crh1p and Crh2p attach chitin to both beta(1-6)- and beta(1-3)glucan in the Saccharomyces cerevisiae cell wall. Eukaryotic Cell 8:1626-1636.
James, T. Y., A. Pelin, L. Bonen, S. Ahrendt, D. Sain, N. Corradi, and J. E. Stajich. 2013. Shared signatures of parasitism and phylogenomics unite Cryptomycota and Microsporidia. Curr Biol 23:1548-1553.
Klis, F. M. 1994. Cell wall assembly in yeast. Yeast 10:851-869.
Kogel-Knabner, I. 2002. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol Biochem 34:139-162.
Kollar, R., B. B. Reinhold, E. Petrakova, H. J. C. Yeh, G. Ashwell, J. Drgonova, J. C. Kapteyn, F. M. Klis, and E. Cabib. 1997. Architecture of the yeast cell wall – beta(1,6)-glucan interconnects mannoprotein, beta(1,3)-glucan, and chitin. J Biol Chem 272:17762-17775.
Latgé, J.-P. 2007. The cell wall: a carbohydrate armour for the fungal cell. Mol Microbiol 66:279-290.
Latge, J.-P. and R. Calderone. 2005. The fungal cell wall. Pages 73-104 in U. Kues and R. Fischer, editors. The Mycota I. Growth, Differentiation and Sexuality. Springer-Verlag, Berlin, Heidelberg.
Shima, J., A. Ando, and H. Takagi. 2008. Possible roles of vacuolar H+-ATPase and mitochondrial function in tolerance to air-drying stress revealed by genome-wide screening of Saccharomyces cerevisiae deletion strains. Yeast 25:179-190.
Treseder, K. K., M. R. Maltz, B. A. Hawkins, N. Fierer, J. E. Stajich, and K. L. McGuire. 2014. Evolutionary histories of soil fungi are reflected in their large-scale biogeography. Ecol Lett 17:1086-1093.
Xie, X. F. and P. N. Lipke. 2010. On the evolution of fungal and yeast cell walls. Yeast 27:479-488.