From: Treseder KK, Lennon JT. 2015. Fungal traits that drive ecosystem dynamics on land. Microbiology and Molecular Biology Reviews 79:243-262.
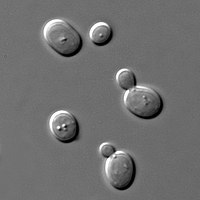
Budding growth forms—typical of yeasts—tend to allow better stress tolerance (Whiteway and Bachewich 2007), perhaps because each cell is encased in a protective cell wall. In contrast, in many filamentous fungi, cells can be connected, allowing water and solutes to flow between them (Cairney 1992, Klein and Paschke 2004). This connectivity can leave the cells more vulnerable to water loss (Beck et al. 2013). Yet, a trade-off of the budding growth form is that single-celled organisms must obtain resources from the micro-environment that immediately surrounds them. Their activities may slow or halt when one or more nutrients become limiting within this micro-site (Schimel and Bennett 2004).
In contrast, filamentous fungi do not have this restriction, since they can forage over relatively long distances—up to several meters for some species (Smith et al. 1992, Legrand et al. 1996, Boddy 1999). As a result, decomposition is often faster when filamentous fungi translocate nutrients to meet their stoichiometric needs—such as transferring N from soil to maintain fungal growth on plant litter with high C:N ratios (Boddy and Watkinson 1995, Frey et al. 2003, Gartner and Cardon 2004, Chigineva et al. 2011, Berglund et al. 2013, Li and Fahey 2013). In this sense, the filamentous growth form can indirectly augment C mineralization in ecosystems, via a mechanism that is not likely to occur with budding growth forms.

http://mmbr.asm.org/content/79/2/243/F1.expansion.html
Lennon et al. (2012) recently reported that fungal taxa differed in preferred moisture availability under laboratory conditions. They assayed yeasts as well as free-living filamentous fungi. In a follow-up analysis of their published data, we observed that the yeasts displayed significantly lower optimum water potentials (i.e., greater drought tolerance) than did free-living filamentous fungi. Other researchers have found that yeasts are common in glacier ice in Antarctica and elsewhere, where water availability and temperature are extremely low (Gunde-Cimerman et al. 2003, Buzzini et al. 2012).

http://mmbr.asm.org/content/79/2/243/F9.expansion.html
Note: Some fungi are dimorphic– they can grow as yeasts or as filamentous fungi. Several of these are pathogens, and will switch to the yeast or filamentous form (whichever is pathogenic) when exposed to the body temperatures of the host. I am not an expert in this area, so I have a question: Have any studies been done on non-pathogenic dimorphic fungi to compare environmental responses for each morphology? In other words, I’d like to know if the yeast form is more stress-tolerant than the filamentous form of the same species. If you know of any, would you mind please leaving a comment?
Beck, J., B. Echtenacher, and F. Ebel. 2013. Woronin bodies, their impact on stress resistance and virulence of the pathogenic mould Aspergillus fumigatus and their anchoring at the septal pore of filamentous Ascomycota. Mol Microbiol 89:857-871.
Berglund, S. L., G. I. Agren, and A. Ekblad. 2013. Carbon and nitrogen transfer in leaf litter mixtures. Soil Biol Biochem 57:341-348.
Boddy, L. 1999. Saprotrophic cord-forming fungi: meeting the challenge of heterogeneous environments. Mycologia 91:13-32.
Boddy, L. and S. C. Watkinson. 1995. Wood decomposition, higher fungi, and their role in nutrient redistribution. Can J Bot 73:S1377-S1383.
Buzzini, P., E. Branda, M. Goretti, and B. Turchetti. 2012. Psychrophilic yeasts from worldwide glacial habitats: diversity, adaptation strategies and biotechnological potential. Fems Microbiology Ecology 82:217-241.
Cairney, J. W. G. 1992. Translocation of solutes in ectomycorrhizal and saprotrophic rhizomorphs. Mycol Res 96:135-141.
Chigineva, N. I., A. V. Aleksandrova, S. Marhan, E. Kandeler, and A. V. Tiunov. 2011. The importance of mycelial connection at the soil-litter interface for nutrient translocation, enzyme activity and litter decomposition. Appl Soil Ecol 51:35-41.
Frey, S. D., J. Six, and E. T. Elliott. 2003. Reciprocal transfer of carbon and nitrogen by decomposer fungi at the soil-litter interface. Soil Biol Biochem 35:1001-1004.
Gartner, T. B. and Z. G. Cardon. 2004. Decomposition dynamics in mixed-species leaf litter. Oikos 104:230-246.
Gunde-Cimerman, N., S. Sonjak, P. Zalar, J. C. Frisvad, B. Diderichsen, and A. Plemenitas. 2003. Extremophilic fungi in arctic ice: a relationship between adaptation to low temperature and water activity. Physics and Chemistry of the Earth 28:1273-1278.
Klein, D. A. and M. W. Paschke. 2004. Filamentous fungi: The indeterminate lifestyle and microbial ecology. Microbial Ecology 47:224-235.
Legrand, P., S. Ghahari, and J. J. Guillaumin. 1996. Occurrence of genets of Armillaria spp in four mountain forests in central France: The colonization strategy of Armillaria ostoyae. New Phytol 133:321-332.
Lennon, J. T., Z. T. Aanderud, B. K. Lehmkuhl, and D. R. Schoolmaster. 2012. Mapping the niche space of soil microorganisms using taxonomy and traits. Ecology 93:1867-1879.
Li, A. and T. J. Fahey. 2013. Nitrogen translocation to fresh litter in northern hardwood forest. Ecosystems 16:521-528.
Schimel, J. P. and J. Bennett. 2004. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 85:591-602.
Smith, M. L., J. N. Bruhn, and J. B. Anderson. 1992. The fungus Armillaria bulbosa is among the largest and oldest living organisms. Nature 356:428-431.
Whiteway, M. and C. Bachewich. 2007. Morphogenesis in Candida albicans. Annual Review of Microbiology 61:529-553.